T-Lymphocytes play a decisive role in the course and clinical outcome of Viral CNS infection.
Summarizing the information presented in this review, the following sequence of events might occur during acute Virus infection.
After invasion of the host and a few initial rounds of replication, the Virus reaches the CNS in most cases by Hematogeneous spread.
After passage through the BBB, CNS cells are infected and replication of Virus in Brain cells causes activation of the surrounding Microglia population.
Moreover, local production of IFN-alpha/beta induces expression of MHC Antigens on CNS cells, and Microglial Cells start to Phagocytose cellular debris, which accumulates as a result of Virus-induced CytoPathogenic effects.
Upon Phagocytosis, Microglia becomes more activated; they up-regulate MHC molecules, acquire Antigen Presentation capabilities and secrete Chemokines.
This will initiate up-regulation of Adhesion Molecules on adjacent Endothelial Cells of the BBB.
Transmigration of activated T-Lymphocytes through the BBB is followed by interaction with APC, presenting the appropriate Peptides in the context of MHC Antigens.
It appears that CD8+ T-Lymphocytes are among the first MonoNuclear Cells to arrive at the infected tissue.
Without a doubt, their induction and attraction is deeply influenced by Natural Killer Cells
Which, after Virus infection, secrete IFN-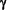 , a Cytokine that stimulates CD8+ T-Cells and diverts the Immune response to a Th1-type CD4+ T-Cell-dominated response.
, a Cytokine that stimulates CD8+ T-Cells and diverts the Immune response to a Th1-type CD4+ T-Cell-dominated response.
Following the CD8+ T-Lymphocytes, tissue-penetrating, Th1 CD4+ T-Cells contact local APC.
This results in a tremendous up-regulation of MHC molecules and secretion of more Chemotactic and toxic substances.
Consequently an increasing number of inflammatory cells, including Macrophages/Microglia and finally AntiBody-secreting Plasma Cells, are attracted to the site of Virus infection.
All trapped cells are mainly terminally differentiated cells that are going to enter Apoptosis during or shortly after exerting their Effector functions.
The clinical consequences and the influence of the effector phase on the further course of the infection depends on the balance and fine-tuning of the contributing Lymphoid Cell populations.
Generally, any delay in the recruitment of Effector Lymphocytes to the tissue or an unbalanced combination of Lymphocyte subsets allows the Virus to spread in the CNS.
Which in turn will cause severe Immune-mediated tissue effects as well as disease.
If either too late or partially deficient, the Immune System response may contribute to a lethal outcome or cause AutoSensitization to Brain-specific Antigens.
By Epitope spreading to the Antigen-Presenting System in Peripheral Lymphoid Tissue.
This could form the basis for subsequent booster reactions of AutoSensitized CD4+ T-Cells - a process that finally will end in an Inflammatory AutoImmune reaction, which in humans we call Multiple Sclerosis.
In contrast, a rapid and specific local response in the Brain tissue will result in efficient limitation of Viral spread and thereby a subclinical Immune System-mediated termination of the infection.
After clearance of Virus-infected cells, downsizing of the local response probably occurs via self-elimination of the contributing T-Cell populations and/or by so far unidentified signal pathways.
However, much of this is highly speculative, and more data have to be collected to make decisive conclusions regarding this matter.
Several strategies have been developed by Viruses to escape T-Cell-mediated eradication.
Including interference with the MHC Class I presentation pathway of the host cell or "hiding" in cells which lack MHC Class I expression.
This may result in life-long persistence of the Virus in the Brain, a state which probably is actively controlled by T-Lymphocytes.
Under severe ImmunoSuppression, however, reactivation of Viral replication can occur, which is a lethal threat to the host.